Biogeography
Biogeography and functioning of microbes at extreme elevations and latitudes.
Our lab has studied microbial life in extreme high elevation ecosystems for the past 15 years, at first focusing on sites in Colorado (e.g. Ley et al. 2004, Schadt et al. 2003) and then expanding to sites in most of the major mountain ranges on Earth (e.g. Darcy et al. 2011, Schmidt et al. 2012). High mountains are perfect environments to study biogeography because they are essentially "islands in the sky" or "outposts of the cryosphere" surrounded by more temperate habitats. As part of this project we also carryout lab and field studies to determine which microbes are actually active in these extreme sites (e.g. Vimercati et al. 2016), and what ecosystem function they are performing (e.g. Schmidt et al. 2009). Research sites are in Colorado, Alaska, Antarctica, Nepal, Chile, Argentina, Tanzania, and Perú. Funding for this work has come from many NSF grants, including two from the now defunct Microbial Observatories Program at NSF (e.g. “The Alpine Microbial Observatory: Microbial Diversity and Function Across Extreme Environmental Gradients”) and several grants from The National Geographic Society (e.g. The Aeolian Zone Revisited: Life in Earth’s Most Extreme Ecosystems).
Our lab has studied microbial life in extreme high elevation ecosystems for the past 15 years, at first focusing on sites in Colorado (e.g. Ley et al. 2004, Schadt et al. 2003) and then expanding to sites in most of the major mountain ranges on Earth (e.g. Darcy et al. 2011, Schmidt et al. 2012). High mountains are perfect environments to study biogeography because they are essentially "islands in the sky" or "outposts of the cryosphere" surrounded by more temperate habitats. As part of this project we also carryout lab and field studies to determine which microbes are actually active in these extreme sites (e.g. Vimercati et al. 2016), and what ecosystem function they are performing (e.g. Schmidt et al. 2009). Research sites are in Colorado, Alaska, Antarctica, Nepal, Chile, Argentina, Tanzania, and Perú. Funding for this work has come from many NSF grants, including two from the now defunct Microbial Observatories Program at NSF (e.g. “The Alpine Microbial Observatory: Microbial Diversity and Function Across Extreme Environmental Gradients”) and several grants from The National Geographic Society (e.g. The Aeolian Zone Revisited: Life in Earth’s Most Extreme Ecosystems).
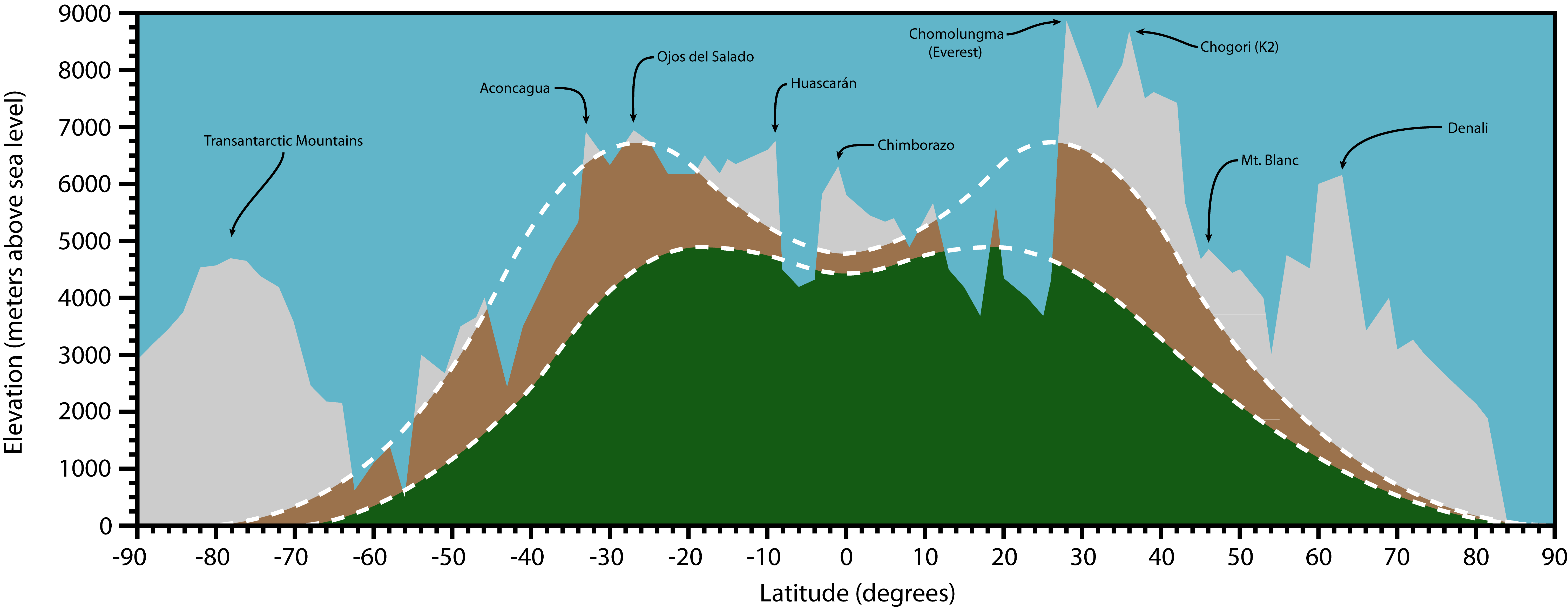
Below: The relationship between latitude and elevation in determining the elevation limits for the mostly plant-free ‘‘cold deserts’’ (brown) environments that we are studying. Vegetated regions are shown in green and permanently ice-covered regions in light white. Negative latitudes represent the Southern Hemisphere. Boundaries between zone are approximate because they can vary dramatically even in one mountain range due to slope and aspect and distance from large bodies of water. The highest mountains in each region are marked for reference. This figure originally appeared here: [Reprint] This figure was created by Jack Darcy.
Relevant publications:
Costello E.K., et al. 2009. Fumarole-supported islands of biodiversity in a hyperarid, high-elevation landscape on Socompa Volcano, Puna de Atacama, Andes. Appl. Environ. Microbiol. 75: 735-747.
Darcy, J.L. et al. 2011. Global distribution of Polaromonas phylotypes - evidence for a highly successful dispersal capacity. PLoS ONE 6: e23742 [Reprint]
Freeman, K.R., A.P. Martin, D. Karki et al. 2009. Evidence that chytrids dominate fungal communities in high-elevation soils. Proc. Natl. Acad. Sci. 106: 18315-18320 [Reprint]
King, A.J. et al. 2010. Biogeography and habitat modelling of high-alpine bacteria. Nature Commun. 1:53 doi: 10.1038/ncomms1055 [Reprint]
Krajick, K. 2006. Living the High Life: The mountaintop environment of the Andes harbors a Noah’s ark of previously undocumented species. Natural Hist. 115: 44-50.
Naff, C.S., J.L. Darcy and S.K. Schmidt. 2013. Phylogeny and biogeography of an uncultured clade of Snow Chytrids. Environmental Microbiology 15: 2672-2680 [Reprint]
Nemergut, D.R., E.K. Costello et al. 2011. Global patterns in the biogeography of bacterial taxa. Environ. Microbiol. 13: 135-144 [Reprint]
Rhodes M., J. et al. 2013. Alpine and Arctic Soil Microbial Communities. in The Prokaryotes (4th ed.), pp. 44-56 (Rosenberg et al., ed.). Springer, Berlin. [Reprint]
Robeson, M.S. et al. 2011. Soil Rotifer communities are extremely diverse globally but spatially autocorrelated locally. Proc. Natl. Acad. Sci. USA 108: 4406-4410 [Reprint]
Schadt, C.W., A.P. Martin, D.A. Lipson and S.K. Schmidt. 2003. Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science 301:1359-1361.
Schmidt S.K. and J.L. Darcy. 2015. Phylogeography of ulotrichalean algae from extreme high-altitude and high-latitude ecosystems. Polar Biology 38: 689-697. [Reprint]
Schmidt, S.K. et al. 2007. Biogeochemical consequences of rapid microbial turnover and seasonal succession in soil. Ecology 88: 1379-1385.
Schmidt S.K. et al. 2009. Microbial activity during extreme freeze-thaw cycles in periglacial soils, 5400 m el., Cordillera Vilcanota, Perú. Extremophiles 13: 807-816.
Schmidt, S.K. et al. 2011. Phylogeography of microbial phototrophs in the dry valleys of the high Himalayas and Antarctica. Proc. Roy Soc. B 278: 702-708 [Reprint]
Costello E.K., et al. 2009. Fumarole-supported islands of biodiversity in a hyperarid, high-elevation landscape on Socompa Volcano, Puna de Atacama, Andes. Appl. Environ. Microbiol. 75: 735-747.
Darcy, J.L. et al. 2011. Global distribution of Polaromonas phylotypes - evidence for a highly successful dispersal capacity. PLoS ONE 6: e23742 [Reprint]
Freeman, K.R., A.P. Martin, D. Karki et al. 2009. Evidence that chytrids dominate fungal communities in high-elevation soils. Proc. Natl. Acad. Sci. 106: 18315-18320 [Reprint]
King, A.J. et al. 2010. Biogeography and habitat modelling of high-alpine bacteria. Nature Commun. 1:53 doi: 10.1038/ncomms1055 [Reprint]
Krajick, K. 2006. Living the High Life: The mountaintop environment of the Andes harbors a Noah’s ark of previously undocumented species. Natural Hist. 115: 44-50.
Naff, C.S., J.L. Darcy and S.K. Schmidt. 2013. Phylogeny and biogeography of an uncultured clade of Snow Chytrids. Environmental Microbiology 15: 2672-2680 [Reprint]
Nemergut, D.R., E.K. Costello et al. 2011. Global patterns in the biogeography of bacterial taxa. Environ. Microbiol. 13: 135-144 [Reprint]
Rhodes M., J. et al. 2013. Alpine and Arctic Soil Microbial Communities. in The Prokaryotes (4th ed.), pp. 44-56 (Rosenberg et al., ed.). Springer, Berlin. [Reprint]
Robeson, M.S. et al. 2011. Soil Rotifer communities are extremely diverse globally but spatially autocorrelated locally. Proc. Natl. Acad. Sci. USA 108: 4406-4410 [Reprint]
Schadt, C.W., A.P. Martin, D.A. Lipson and S.K. Schmidt. 2003. Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science 301:1359-1361.
Schmidt S.K. and J.L. Darcy. 2015. Phylogeography of ulotrichalean algae from extreme high-altitude and high-latitude ecosystems. Polar Biology 38: 689-697. [Reprint]
Schmidt, S.K. et al. 2007. Biogeochemical consequences of rapid microbial turnover and seasonal succession in soil. Ecology 88: 1379-1385.
Schmidt S.K. et al. 2009. Microbial activity during extreme freeze-thaw cycles in periglacial soils, 5400 m el., Cordillera Vilcanota, Perú. Extremophiles 13: 807-816.
Schmidt, S.K. et al. 2011. Phylogeography of microbial phototrophs in the dry valleys of the high Himalayas and Antarctica. Proc. Roy Soc. B 278: 702-708 [Reprint]
